What determines pollen grain shape? One might reasonably assume that dispersal method (also called pollination syndrome) is the primary driver of pollen morphology. Indeed, research has shown that wind-dispersed pollen tends to be smoother and smaller than insect-dispersed pollen. The latter, in contrast, is usually more complex in shape, often displaying spikes or hooks on its surface. On the other hand, evolutionary theory suggests that the pollen morphospace (the range of allowable shapes a pollen grain can take) is tightly constrained by phylogeny. In other words: closely related species should display similar pollen shapes.
What happens, then, when closely related species produce pollen that are dispersed by different mechanisms? This is precisely the question I sought to answer in this mini-project. All subsequent work was conducted at the Jasper Ridge Biological Preserve (Ootchamin 'Ooyakma) as part of the BIO 105 docent-training course.


Left: common yarrow (Achillea millefolium), an insect-dispersed pollen producer. Image from Prides Corner Farm. Right: California sagebrush (Artemisia californica), a wind-dispersed species endemic to California. Image from Wikipedia.
California sagebrush (Artemisia californica) is unique among the Asteraceae's Anthemida tribe members, which tend to be insect-pollinated. Perhaps the wind-dispersal mechanism of sagebrush pollen reflects an adaptation to its late summer flowering window, when pollinators are less abundant. Yarrow and sagebrush belong to distinct genera, but share a recent common ancestor (estimated ~20-30mya) and generally inhabit similar environments. At Jasper Ridge, both species can be found on the edge of grassland trails. We hypothesize that despite their shared evolutionary history, sagebrush and yarrow pollen will display distinct morphologies tuned to their respective method of dispersal: smooth and simple for wind-dipsersed sagebrush, complex and spiny for insect-dispersed yarrow. While previous surveys of pollen morphology have been conducted in the Asteraceae family, comparisons between closely related species with distinct pollination syndromes remain rare.
Pollen grains were collected directly from the field (for yarrow) or from herbaria specimen in the Oakmead collection at Jasper Ridge (for sagebrush). Grains were gold-sputtered and mounted for imaging following standard SEM sample prep workflow. Grains were imaged on JEOL JCM-7000 NeoScope SEM at 900× magnification and 10 kV accelerating voltage.
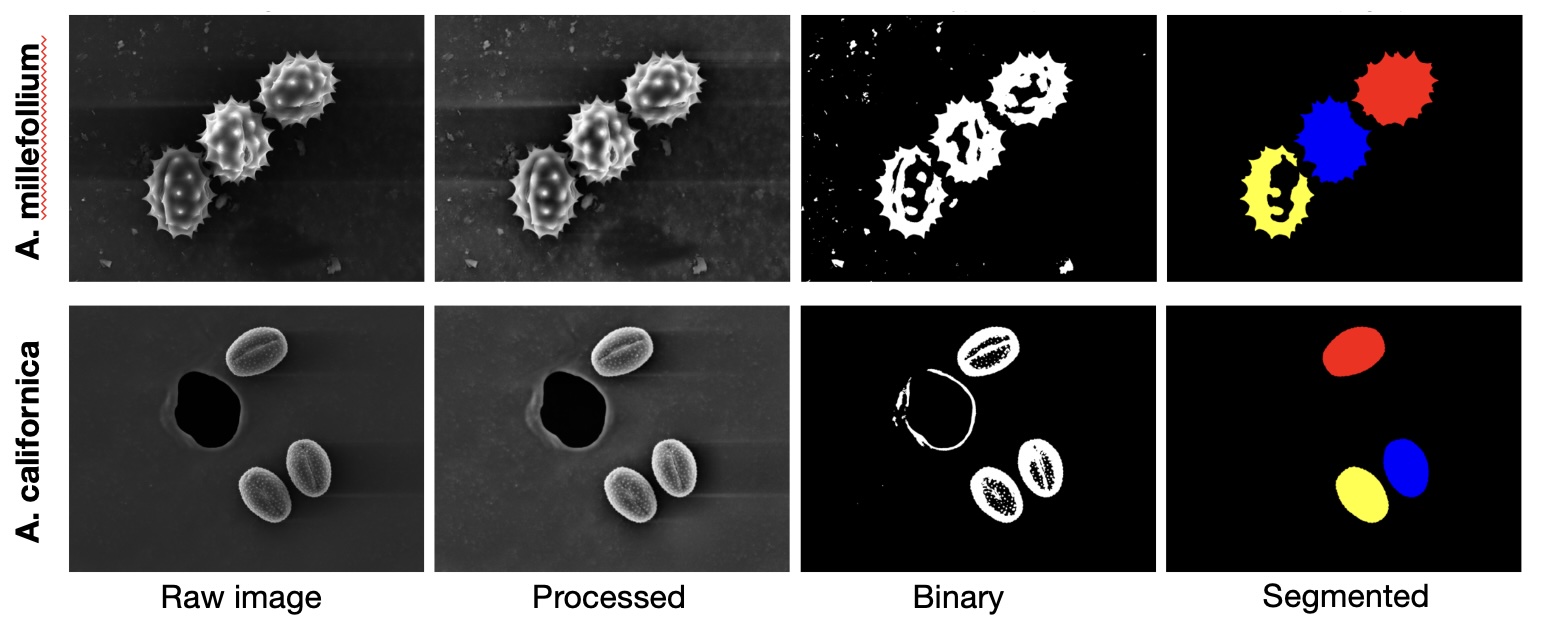
Sample images from image segmentation pipeline. Code is available upon request.
A custom Python-based image processing and segmentation pipeline was created for this project. Segmented regions were retained as individual pollen grains if they satisfied the following criteria: area 30,000–350,000 px², circularity ≥ 0.20, aspect ratio ≤ 1.8, and centroid located ≥ 40 px from the image border. In total, 50 yarrow grains and 51 sagebrush grains were retained. For each retained grain, the following morphometric variables were extracted: diameter, area, perimeter, aspect ratio, circularity, solidity, spine count, spine density and roughness range. All linear and area measurements were converted to physical units using the instrument pixel size.
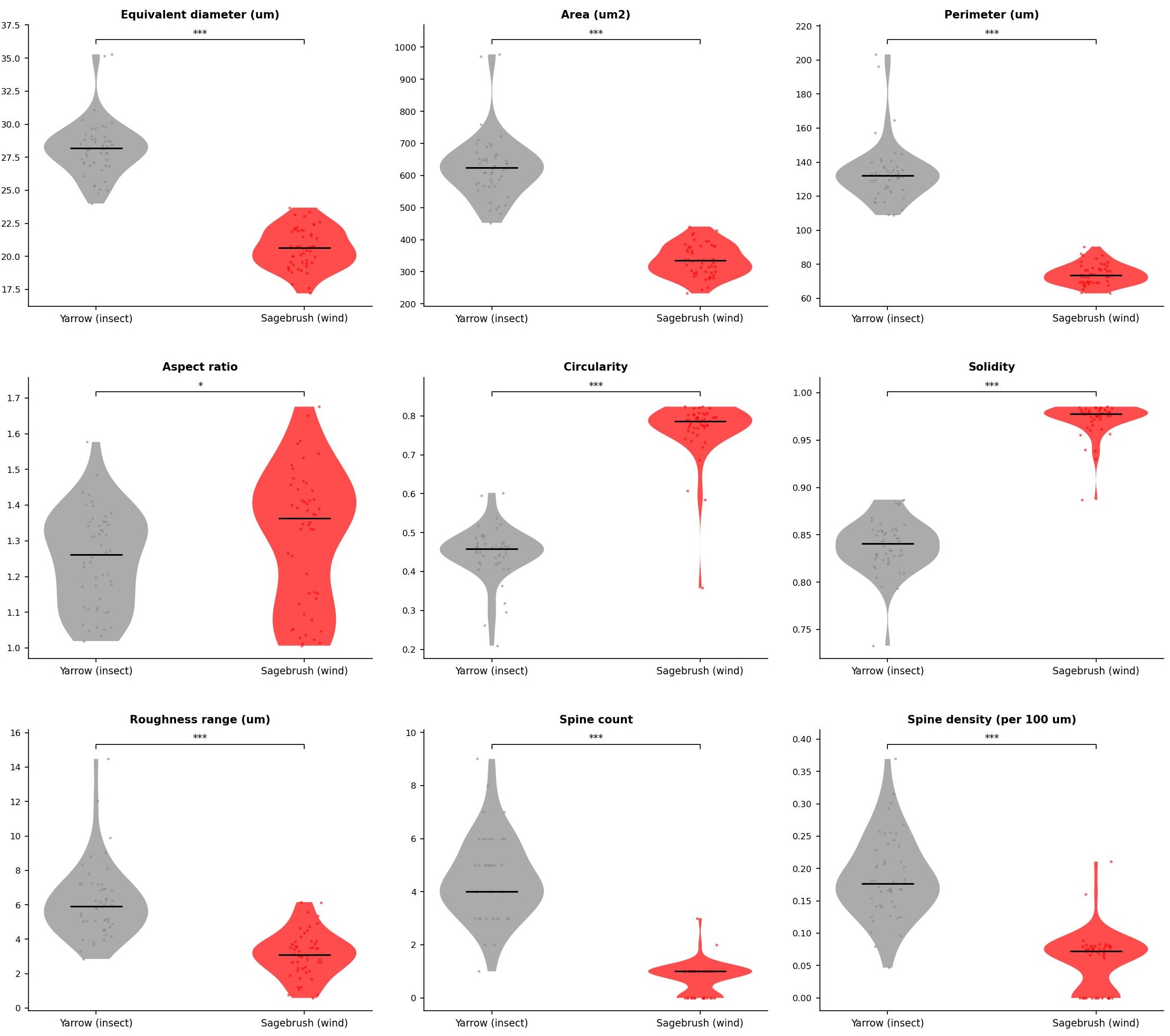
Pollen grain morphometric measurements. Pairwise statistical testing by Mann-Whitney U tests (*: p < 0.05; **: p < 0.01; ***: p < 0.001). Analysis code available upon request.
Quantitative comparisons between pollen grains supported the prediction that pollination syndrome shapes grain architecture, even within a plant tribe. Yarrow grains are consistently larger (by up to 37%) and spinier than those of sagebrush , and this distinction was captured robustly across the nine variables tested. Larger grain size in entomophilous species is a well-documented trend and is thought to reflect the reduced energetic cost of biotic transport: insect-mediated delivery does not impose the same aerodynamic size penalty as wind dispersal, relaxing selection for miniaturization.
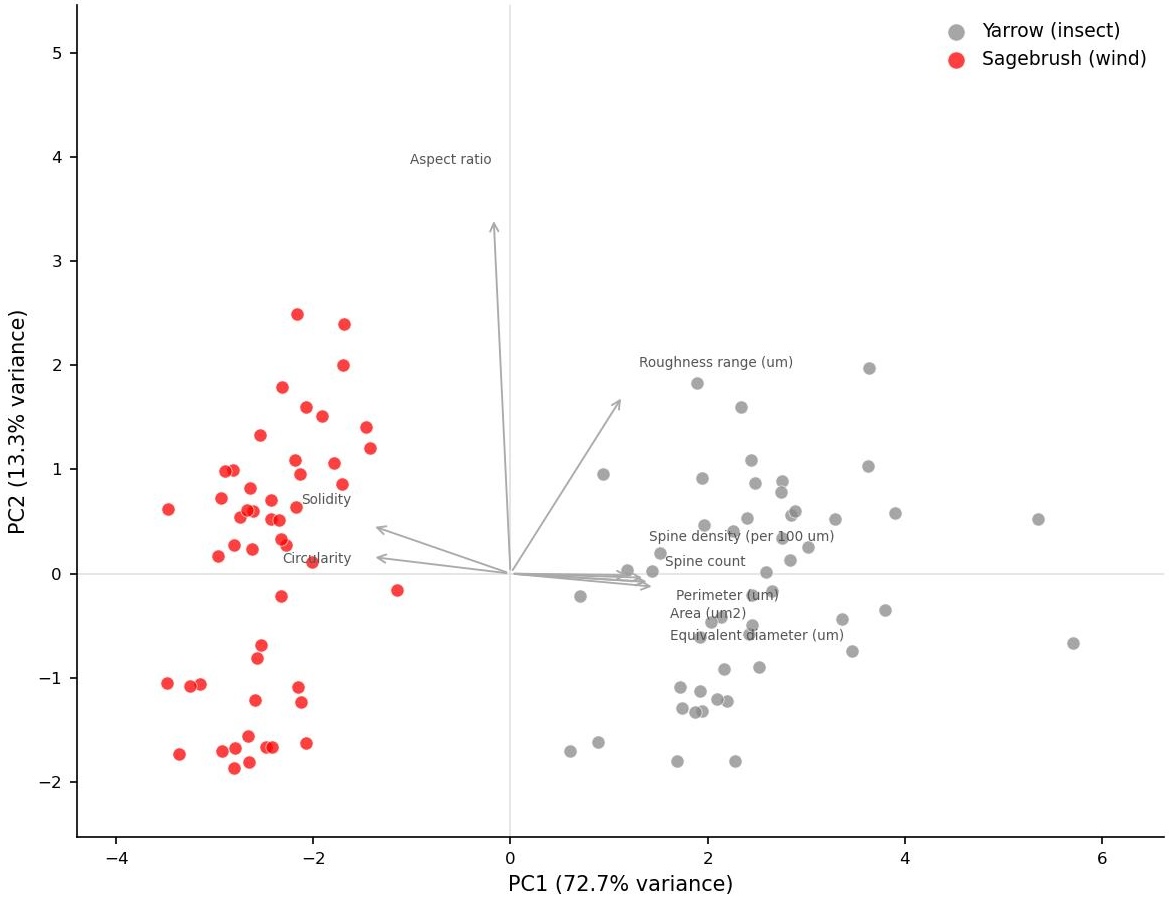
Principal component analysis of pollen morphology. Code available upon request.
The two species occupied largely non-overlapping regions of pollen morphospace in the PCA, with a single PC cleanly separating them. These results are consistent with aerodynamic and mechanical frameworks for pollen evolution: wind-dispersed pollen is predicted to be small, smooth, and spherical to minimize drag and maximize suspension time in air, while insect-dispersed pollen is predicted to be larger and ornamented with structures that facilitate adhesion to pollinator body hair.
There are important limitations to this study: the sample includes only one representative species per pollination strategy. A stronger comparison would include more closely related species, for example two species in the same genus that exhibit distinct methods of grain dispersal. What we can conclude for now is that pollination syndrome is a strong predictor of pollen grain architecture even within a single plant tribe and that automated image analysis pipelines can reliably recover biologically meaningful morphological signal from SEM micrographs.
Special thanks to Shan-Wei Sun, Jack Riley, Adriana Hernandez, Katie Glover and Rodolfo Dirzo, without whom this project would not have been possible.
References
- Timerman D, Barrett SCH. (2021). The biomechanics of pollen release: new perspectives on the evolution of wind pollination in angiosperms. Biol Rev 96(5):2146–2163.
- Jardine PE et al. (2022). Why does pollen morphology vary? Evolutionary dynamics and morphospace occupation in the largest angiosperm order (Asterales). New Phytologist 234(3):1075–1087.
- Lu L-L et al. (2022). Artemisia pollen dataset for exploring the potential ecological indicators in deep time. Earth Syst Sci Data 14:3961–3995.
- Faegri K, van der Pijl L. (1979). Principles of Pollination Ecology, 3rd ed. Pergamon Press, Oxford.